简单总结:采用疫苗免疫措施主要用于预防猪生殖和呼吸综合征(PRRS)。我们总结了目前开发的PRRSV疫苗的优缺点和适用性,包括修饰活病毒、灭活病毒、重组亚基、活载体、DNA、基因缺失、合成肽、病毒样颗粒疫苗,以及其他各种疫苗。这些结果为新疫苗的开发提供了理论基础。
摘要:猪繁殖与呼吸综合征(PRRS)是养猪业中的一种高度传染性疾病,但其发病机制尚不完全清楚。这种疾病是由猪呼吸综合征病毒(PRRSV)引起的,它主要感染猪肺泡巨噬细胞并破坏免疫系统。不幸的是,目前还没有治疗PRRS的特效药物,因此疫苗接种对于控制该疾病至关重要。现有多种类型的单一疫苗和联合疫苗,包括活疫苗、灭活疫苗、亚基疫苗、DNA疫苗和载体疫苗。其中,活疫苗具有较好的保护作用,但交叉保护作用较弱。灭活疫苗较为安全,但免疫效力较差。亚单位疫苗可用于妊娠晚期,DNA疫苗可增强活疫苗的保护作用。然而,载体疫苗只提供部分保护,并没有在实践中广泛使用,目前仍然需要一种符合新一代国际标准的PRRS疫苗。本文全面综述了减毒活疫苗、灭活疫苗、亚基疫苗、活载体疫苗、DNA疫苗、基因缺失疫苗、合成肽疫苗、病毒样颗粒疫苗以及其他预防和控制PRRS的疫苗的优缺点和适用性。其目的是为疫苗研究和开发提供理论基础。
关键词:猪繁殖与呼吸综合征;疫苗;优点和缺点;适用性;研究进展。
1、引言
猪繁殖与呼吸综合征(PRRS)是由猪繁殖与呼吸综合征病毒(PRRSV)引起的一种高传染性和强毒性传染病。可引起不同年龄猪的流产、早产、胎儿干尸等生殖障碍及呼吸系统疾病。猪感染PRRSV可导致免疫抑制,损害免疫功能,给养猪业造成重大经济损失。PRRS于1987年首次在北卡罗来纳州被发现,此后在世界范围内分离出各种毒株。1991年和1992年,在欧洲和美洲的感染猪中分离出Lelystat病毒和VR2332株。PRRSV LV和VR2332株遗传分类为1型PRRSV (PRRSV-1)和2型PRRSV (PRRSV-2)。在最近的系统分类中,PRRSV-1和PRRSV-2分别归属于乙型肝炎病毒1和乙型肝炎病毒2种。PRRSV-2毒株主要流行于中国,那里曾发生高致病性PRRS (HP-PRRS)疫情并伴随仔猪的高死亡率。2012年,中国发现了一株高度可变的NADC30样毒株,该毒株极易与其他毒株重组。此外,NADC30样PRRSV在中国许多地区流行。PRRSV继续变异和重组,目前的疫苗不能提供足够的保护,使预防和控制更加复杂。
PRRSV是一种带包膜的单正链RNA病毒,属于动脉病毒属。其基因组长度约为15 kb,由ORF1a、ORF1b、ORF2a、ORF2b、ORF3-7、ORF5a和ORF1aTF等11个开放阅读框(ORFs)组成,与ORF1a的非结构蛋白(NSP)- 2编码区重叠。其中ORF1a、ORF1aTF和ORF1b占总基因组的80%。翻译后的多肽pp1a和pp1ab被切割成15个非结构蛋白(NSPs),包括NSP1α、NSP1β、NSP2相关蛋白(NSP2N、NSP2TF和NSP2)和NSP3-NSP12,它们参与病毒复制。ORF2a、ORF2b、ORF3、ORF4、ORF5和ORF5a编码6个包膜相关蛋白,分别为GP2a、GP2b、GP3、GP4、GP5和GP5a。ORF6编码的非糖基化基质(M)蛋白和ORF7编码的核衣壳(N)蛋白是优势结构蛋白,具有较强的免疫原性。GP5蛋白是一种主要的结构蛋白,具有良好的免疫原性,能够诱导机体产生中和抗体(neutralizing antibodies, NAs),在持续病毒感染中起着至关重要的作用。因此,GP5仍是PRRS疫苗研究的首选蛋白。
抗体依赖性增强(ADE)是PRRS疫苗开发的一个重大挑战,其中先前存在的非中和或亚中和抗体促进病毒进入和复制。首例PRRSV感染的ADE的报道可追溯到1993年。一项研究发现,与仅接种病毒的胎儿相比,接种病毒加抗体的胎儿的病毒复制增加。Yoon等人后来报道,在病毒感染前给猪注射亚中性量的PRRSV特异性免疫球蛋白G(IgG)会增加病毒血症,并且PRRSV特异性IgG会增加病毒逃逸突变体的产生。Fcγ受体可参与PRRSV引起的ADE。然而,有研究发现亚中和的IgG在体内不会引起ADE,经修饰的PRRSV活疫苗可对异源毒株提供部分交叉保护。目前,体内实验还不足以证实ADE是否在PRRSV发病中起关键作用。感染PRRSV后的ADE可能因不同的毒株和条件而异。体外研究表明,抗PRRSV血清在稀释27倍时对感染具有最有效的作用。ADE试验采用1头样品与阳性对照比为1.2的猪的血清,其NA效价为1/5.9。在PRRSV NA被动转移后,血清中较高浓度的PRRSV NA滴度(1:32)可诱导完全保护。然而,这只适用于一些仔猪,并且滴度(1:8)不能阻止PRRSV在肺部复制或传播到其他周围淋巴组织。这些研究结果是相互矛盾的,需要进一步研究确定PRRSV感染期间ADE的潜在机制。确定减毒PRRS疫苗是否会使感染新毒株的猪的疾病恶化也是必要的。
目前,疫苗接种是预防PRRS的主要措施。国产灭活疫苗来源于CH-1a经典毒株,而应用广泛的减毒疫苗来源不同,分为经典毒株(VR2332、CH-1R、R98)和高致病性毒株(JXA1、TJ、HuN4、GD)。中国流行的PRRSV可分为4个谱系,即谱系1、谱系3、谱系5和谱系8。自PRRSV在中国出现以来,谱系8占主导地位,包括2006年之前流行的经典PRRSV毒株(CH-1a-like)和2006年之后流行的HP- PRRSV-like毒株。谱系1也被称为NADC30-like毒株,自2013年以来在全国范围内迅速传播,目前的临床检出率与谱系8(HP- PRRSV-like) 相当。虽然谱系5 (BJ-4-like/VR2332-like)早在1996年就出现了,但在中国一直处于非大流行状态,临床检出率较低。
与谱系8(CH-1a-like)和谱系5相比,HP-PRRSV-like和NADC30- like毒株表现出更高的遗传变异和重组率。这些特征可能使现有疫苗失效,使其更容易逃避免疫监视。此外,目前的PRRS疫苗的局限性超出了它们对异源毒株的保护能力。它们也不能满足养猪业对全面和普遍保护的需求,并有恢复毒性的风险。尽管商业疫苗在病毒学和临床环境中对同源毒株提供了合理的保护,但它们并非万无一失。继续研究和开发基因工程疫苗和标记疫苗可以解决目前PRRS疫苗的局限性。本文综述了各种PRRS疫苗的优点、缺点和适用性,包括改良活病毒(MLV)疫苗、灭活疫苗、亚单位疫苗、活载体疫苗、DNA疫苗、基因缺失疫苗、合成肽疫苗、病毒样颗粒疫苗(VLP)疫苗以及其他各种疫苗。目标是为开发可以在受该疾病影响的地区增强对PRRSV的保护的新疫苗提供理论基础。在下一代疫苗开发的背景下,进一步评价蛋白质组全结构域肽库的筛选方法和鉴定出的抗原,将对开发出能够更好地在中国预防PRRSV的新疫苗,特别是针对异源毒株和遗传变异率和重组率较高的毒株具有重要作用。
2、MLV疫苗
自发现PRRSV以来,已经开发了几种改良活病毒(MLV)疫苗来对抗PRRSV-1和PRRSV-2。针对PRRS1-1开发的Porcilis PRRS (Merck, Madison, NJ, USA)、UNISTRAIN PRRS (Hipra, S.A, Amer, Spain)、Ingelvac PRRSFLEX EU (Boehringer Ingelheim, Ingelheim am Rhein, Germany)、ReproCyc PRRS EU (Boehringer Ingelheim, Ingelheim am Rhein, Germany)和Suvaxyn PRRS MLV (Zoetis, Belgium, WI, USA)分别于2000年、2013年、2015年、2015年和2018年获批上市,在西欧国家广泛使用。针对PRRSV-2开发的Ingelvac PRRS MLV (Boehringer Ingelheim, Ingelheim am Rhein, Germany)、Fostera PRRS (Zoetis, Belgium, WI, USA)、PrimePac PRRS (Merck, Madison, NJ, USA)和prevent PRRS (Eli Lilly, Ind, USA)疫苗分别于1994年、2012年、2014年和2018年获批上市,主要在美国使用。疫苗的功效和疫苗引起的免疫详情见表1。
表1.国外MLV疫苗统计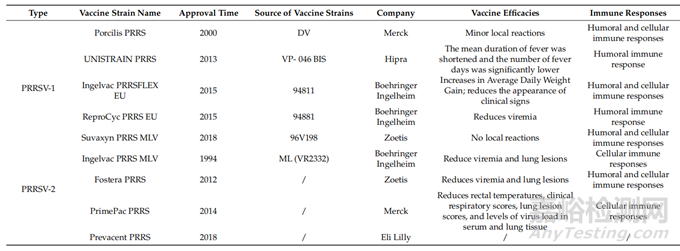
在中国,主要是针对PRRSV-2开发的PRRS疫苗,MLV疫苗有Ingelvac PRRS、CH-1R、R98、JXA1-R、TJM-F92、HuN4-F1、GDr-180等7种(表2),但没有一种疫苗可以声称完全预防该病。PRRSV,无论是野生型还是减毒型,都会诱导低水平的细胞免疫,并且直到感染后期才产生核NAs。研究表明,接种疫苗后,猪出现病毒血症,但无法检测到NAs水平,病毒特异性γ-干扰素分泌细胞(IFN-γ-SC)频率降低。外周血单核细胞(PBMCs)产生的白细胞介素-10(IL-10)水平与IFN-γ反应呈负相关。这些结果表明,IFN-γ和IL-10在对PRRSV免疫的发展中起重要作用。MLV对猪的保护作用不是基于体液因素,而是基于细胞介导的免疫。总体而言,现有证据表明,所有先前批准的PRRS MLV疫苗都会引起较弱的体液和细胞免疫反应。
表2.中国疫苗统计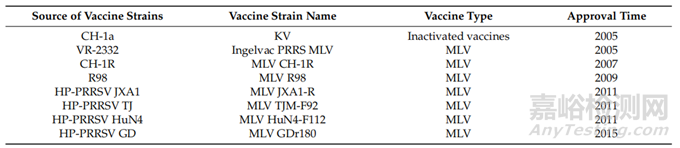
评估疫苗效力的实验表明,猪呼吸与繁殖综合征减毒活疫苗(PRRSV- MLVs)对基因同源的野生型PRRSV毒株具有足够的保护作用,但对异源型PRRSV毒株仅提供部分保护或没有保护作用。2012年,研究表明,与未接种MLV疫苗的猪相比,接种疫苗的猪死亡率显著降低,发烧天数减少,卡他性支气管肺炎发病率降低,体重增长增加,病毒血症水平降低。该研究发现,用减毒的PRRSV-1疫苗免疫可以部分保护机体免受高毒力的PRRSV-2毒株的攻击。Bonckaert等在另一项检测商业化减毒PRRSV-1亚型活疫苗UNISTRAIN有效性的实验中发现,接种后猪的平均发热时间缩短,发热天数明显低于对照猪。少数接种疫苗的动物出现发烧、呼吸系统疾病和结膜炎。疫苗接种组刺激后鼻分泌物病毒平均滴度显著低于对照组。与对照组相比,疫苗组病毒血症持续时间略有减少,但差异无统计学意义。最近的研究表明,针对PRRSV-1的MLV疫苗可以保护1日龄仔猪在断奶时免受异源PRRSV-1感染。此外,针对PRRSV-2的MLV疫苗已被证明在妊娠母猪的整个孕期对异源PRRSV-1具有交叉保护作用。它们还对HP-PRRSV感染具有保护作用。然而,在妊娠晚期,针对PRRSV-1的MLV疫苗只能保护后备母猪免受异源PRRSV-1的侵害,而不能保护后备母猪免受PRRSV-2的侵害。Maragkakis等人进行了一项研究,他们观察到与肌肉注射相比,仔猪皮内注射PRRSV-1 MLV疫苗对其健康和生产性能有有益的影响。这种积极的影响可以通过屠宰时体重的增加和肺和胸膜炎病变评分的降低来证明。因此,目前所有的MLVs都不能提供全面的保护,不能完全满足养猪业的需求。
因为垂直传播、出生仔猪存活数量减少以及可能在接种疫苗的宿主中引起持续感染,PRRSV-MLV的安全性面临挑战。Charerntantanakul和Wang等人证明,接种MLV疫苗的猪在免疫后可出现长达四周的病毒血症,导致疫苗病毒传播给未接种疫苗的猪。因此,应考虑PRRSV-MLV的安全性。自1994年Ingelvac PRRS MLV疫苗获准上市以来,该疫苗已在中国和美国的养猪场被广泛用于疫苗接种,并发现导致这些养猪场随后爆发PRRS的病毒与两国报告的疫苗株具有几乎相同的核苷酸序列。此外,MLVs与野生型毒株之间的重组也加剧了PRRS的爆发。最近的研究表明,丹麦的一种新毒株是由涉及Amervac株(单株PRRS疫苗;Hipra)和96V198株(Suvaxyn PRRS;Zoetis AH)。96V198菌株主要贡献了该菌株的基因组成,包括ORFs1 -2和部分ORF3。另一方面,Amervac菌株作为次要亲本,贡献了基因组的剩余部分。因此,对新型PRRSV-MLV的研究将在疫苗的有效性和安全性方面取得突破。
在临床上,MLV疫苗通常用于已经爆发过PRRS的农场。对于没有发生PRRS的养殖场,MLV疫苗通常用于预防疾病,而不是用于预防感染。接种疫苗后,疫苗中的减毒株会在猪体内繁殖,刺激机体产生抗体来对抗疾病。MLV对3-18周龄的猪具有事宜的免疫预防作用。此外,疫苗接种后7天内即可发挥保护作用,保护期较长,可达16周以上。母猪或替代后备母猪的免疫接种也能提高其繁殖性能。疫苗毒株决定减毒疫苗的有效性,PRRSV基因多样性影响减毒疫苗的有效性。此外,该疫苗对同源毒株具有良好的预防作用。MLVs对预防PRRS有特殊作用;然而,由于潜在的安全问题,在使用前应仔细选择疫苗株。
3、灭活疫苗
与MLV疫苗相比,PRRSV灭活疫苗由于安全性提高,已在全球范围内获得许可。在中国和其他国家有许多针对PRRSV-2株(CH-1a、ISU-P、JXA1、VD-A1和SD-1)和PRRSV-1株(LV、VD-E1、VD-E2和PRRS-CY-218-JPD-P5-6-91或Olot)的市售灭活疫苗。然而,自2005年以来,由于观察到这些疫苗的疗效较差,美国已不再提供这些疫苗。Opriessnig等人证实,PRRSV灭活疫苗可缩短感染后病毒在公猪精液中的持续时间。然而,Nilubol在2年后用同一来源的灭活疫苗(PRRomisee、Intervir、VR-2402疫苗株)对公猪进行免疫,但未能显示精液中病毒含量的减少。两项试验的不同之处是,前一项研究使用的感染株与疫苗株(VR-2402)相同,后一项研究使用的感染株为VR-2385,这说明灭活疫苗诱导的交叉保护较弱。与此同时,一些报道阐明了PRRSV灭活疫苗对野生病毒感染的不良影响,其表现为缺乏可检测到的PRRSV特异性抗体产生和细胞介导的免疫(CMI)应答。在两种基因型PRRSV灭活疫苗诱导的PRRSV特异性免疫应答(KV/ADJ、Progressis、Merial Labs、PRRSV-1和PRRomisee、Intervet、PRRSV-2)中,病毒NAs的滴度一般低于8,不能有效清除PRRSV。因此,在体内实验中,PRRSV灭活疫苗在感染野生型PRRSV的猪之间没有产生统计学差异。
PRRSV灭活疫苗具有安全、储存方便、便于运输等优点,一般情况下,母源抗体对PRRSV灭活疫苗的干扰较弱。此外,接种灭活疫苗的母猪死胎数和流产率明显降低,母猪的繁殖力和仔猪存活率均有提高。然而,灭活疫苗也有一定的缺点,如对异源毒株的免疫效果差,接种剂量大,需多次免疫,成本高,产生免疫需要特定的一段时间。国内分离的病毒株(CH-1a株)制成的灭活油佐剂疫苗具有良好的免疫调节作用,且维持时间较长。第二次免疫后仅20天猪即能获得免疫,这是至关重要的。它还为80%以上接种过疫苗的猪提供了良好的保护,并可提供长达6个月的免疫期。野生型病毒株(SI株)灭活油佐剂疫苗适用于仔猪免疫。一旦动物接种疫苗,它可以提供超过6个月的保护。同时,该疫苗还可用于替代后备母猪和公猪的接种。最近的研究证实了匈牙利成功实施了PRRS根除规划。这一成就是通过给母猪注射灭活疫苗和对其后代采用隔离饲养来实现的。值得注意的是,在整个种群更替过程中,生产几乎保持不间断。一些研究表明,使用MLV疫苗和灭活疫苗的组合可以增强猪对不被PRRSV感染的保护。在母猪妊娠第8周接种MLV疫苗,然后在分娩前3周再次接种商业灭活疫苗,可有效降低PRRSV的发生率。并且,有研究指出,断奶仔猪PRRSV血清阳性个体的比例更高。此外,MLV疫苗与灭活疫苗联合使用可对仔猪提供部分保护,减少病理性肺损失,改善体重增加,降低病毒血症。这种影响可能归因于灭活疫苗诱导母猪产生NAs。总之,这些数据表明,灭活PRRSV疫苗可能在治疗PRRSV方面具有潜在的治疗作用,而不是预防疾病。因此,在开发含有佐剂的新配方之前,可能需要进一步探索灭活疫苗的治疗益处。
4、亚单位疫苗
亚单位疫苗又称生物合成亚单位疫苗,是将特异性抗原编码基因插入易增殖的病毒或细胞中,表达有效的特异性抗原并对其进行纯化而制成的。虽然它们是安全稳定的,但它们的免疫原性通常很低。免疫原性的PRRSV蛋白以结构和非结构形式存在。该病毒有六种膜蛋白,标记为主要((GP5和M)和次要(GP2a、E、GP3和GP4)包膜蛋白。GP5是最丰富的糖蛋白,也是NAs的主要诱导剂,但在PRRSV毒株间差异较大。M蛋白是一种高度保守的非糖基化的结构蛋白,有助于病毒的组装和出芽。在感染病毒的细胞中,GP5和M存在于由共价二硫键连接的“异二聚体复合物”中。N蛋白在病毒感染细胞中最为丰富,约占病毒粒子的40%。次要包膜蛋白通过非共价相互作用在感染细胞中形成“异四聚体复合物”。除了GP5,次要蛋白也具有病毒中和的B细胞抗原表位。PRRSV的潜在T细胞表位存在于蛋白GP3、GP4、GP5、M、N、NSP2、NSP5、NSP9和NSP10中。为了诱导有效的抗PRRSV免疫,在创新疫苗配方中以“异二聚体和异四聚体复合物”的形式暴露免疫原性病毒蛋白,以有效诱导特异性和保护性B细胞和T细胞反应是至关重要的。在PRRSV发现后不久,杆状病毒表达的PRRSV结构蛋白被作为潜在的亚单位疫苗进行了测试。在最早的研究中,Plana等通过构建PRRSV ORF2-ORF7表达载体进行了动物免疫试验。结果显示GP3和GP5具有特异性保护作用,这表明GP3和GP5亚单位疫苗具有良好的免疫原性,可作为开发重组亚单位疫苗的候选基因。此后,也测试了一种抗PRRSV的转基因植物口服亚单位疫苗。然而,基于植物的实验性疫苗与基于杆状病毒的亚单位疫苗具有相同的缺陷;具体而言,其对猪的功效有限。Song等通过重叠PCR扩增出缺失信号肽和跨膜序列的GP5基因截断片段,将其插入原核表达载体pET-32a或pGEX-6p-1中,添加His或GST标签诱导表达和纯化,并进行动物实验。结果表明,鼠伤寒沙门菌鞭毛蛋白是一种有效的佐剂,因为它可以增强抗GP5抗体的诱导,并诱导体液和细胞免疫反应。然而,本研究是在小鼠模型上进行动物免疫实验,该疫苗在猪身上的效果尚不完全清楚。转基因玉米愈伤组织产生PRRSV M蛋白,并通过口服转基因植物组织诱导小鼠血清和肠黏膜抗原特异性抗体。结果表明,转基因玉米是生产亚单位疫苗的有效途径,可产生对PRRSV的全身和粘膜免疫应答。An等]在拟南芥植物中成功表达了PRRSV衍生的抗原蛋白,实现了重组蛋白疫苗的成功生产。Peng等比较了四种天然佐剂的免疫增强作用以提高PRRSV亚单位疫苗的效力,结果发现黄芪/芽孢杆菌佐剂对PRRSV GP5亚单位的免疫增强作用最为有效。Peng等也研究了泰山马尾松花粉多糖(TPPPS)和Freund佐剂对PRRSV亚单位疫苗的免疫增强作用。基于上述结果,预计中等剂量的TPPPS作为GP5佐剂可成为候选PRRSV亚单位疫苗。
与传统疫苗相比,亚单位疫苗不含核酸物质;因此,它们更安全。动物免疫不会导致持续感染,故其免疫反应可与野生病毒感染的免疫反应区别开来。一些研究表明,在妊娠晚期接种PRRSV亚单位疫苗可有效预防由异种PRRSV-1和PRRSV-2感染引起的生殖失败。这有利于预防和控制各种疾病。但其生产成本较高,免疫原性可能优于减毒疫苗或灭活疫苗;因此,它的发展时间相对较长。从长远来看,PRRSV亚单位疫苗在预防和控制PRRSV方面具有广阔的应用前景。然而,其免疫持续时间短,必须与佐剂联合使用。佐剂通过增强参与抗原免疫应答的巨噬细胞的活性来促进T细胞或B细胞的应答。研究发现多种细胞因子可增强疫苗的特异性免疫应答;因此,开发具有有益作用的佐剂将有助于增强PRRSV亚单位疫苗的特异性免疫应答。
5、活载体疫苗
利用基因工程技术将重组PRRSV抗原基因转化为能够表达外源病毒蛋白的活载体,制备了PRRSV活载体疫苗。该疫苗经动物直接免疫后可引起特异性免疫应答[83]。PRRSV活载体疫苗主要以痘病毒、腺病毒、疱疹病毒和猪伪狂犬病毒(PRV)为载体表达PRRSV中心免疫原性基因。Alonso等成功构建了表达GP5蛋白的重组传染性胃肠炎病毒,并检测到高水平的GP5特异性抗体。Fang等构建了表达PRRSV GP5和M蛋白的重组PRV载体。结果表明,重组病毒载体疫苗能诱导NA的产生,并能刺激淋巴细胞增殖。Tian等以PRRSV (DS722株)为活病毒载体,构建了表达猪流感病毒(SIV)和猪圆环病毒2型(PCV-2)保护性抗原的多组分疫苗病毒DS722-SIV- PCV2。实验结果表明,DS722-SIV-PCV2具有作为PRRSV三价候选疫苗的潜力,同时也揭示了其作为PRRSV活疫苗载体的可能性。近年来的研究表明,表达猪瘟病毒E2蛋白的重组PRRSV载体的候选疫苗rPRRSV-E2是有效预防和控制HP-PRRSV和猪瘟的必要条件。近年来,活病毒载体在PRRSV疫苗研究中显示出巨大的潜力,但其生产成本高,效率低,且载体病毒也可能引发免疫反应,从而降低疫苗的有效性。尽管研究人员仍然不了解病毒的自然发生,但人们对活病毒载体疫苗株与野生或减毒病毒疫苗株之间是否会发生重组存在疑问。尽管如此,我们相信随着生物技术的发展,活病毒载体疫苗有可能成为PRRSV疫苗领域的焦点。
6、DNA疫苗
DNA疫苗又称核酸或基因疫苗,其原理是将产生病原微生物特异性抗原的真核表达载体直接引入动物体内,刺激产生典型的免疫反应。Kwang等构建了PRRSV ORF4-ORF7四种重组DNA疫苗,并进行了动物免疫试验。通过ELISA、病毒中和和Western blotting分析,71%的免疫动物检测到PRRSV特异性抗体。此外,通过IFN-γ和/或增殖试验,86%的免疫猪检测到细胞免疫应答。结果证实PRRSV抗原中和表位主要集中在GP4和GP5蛋白上,表明DNA疫苗可诱导体液免疫和细胞免疫。Siriseewan等人发现,在MLV免疫前2周使用编码截断PRRSV N蛋白的DNA疫苗进行预免疫,可以提高PRRSV特异性免疫,这可以通过增加NA滴度和PRRSV特异性IFN-γ的产生以及减少IL-10和PRRSV特异性Treg的产生来观察。用PRRSV GP5嵌合序列构建的DNA疫苗也与阳离子脂质体络合,并皮下和肌肉注射到实验猪体内。结果显示,接种GP5-Mosaic疫苗的猪外周血单个核细胞中IFN-γ mRNA的表达量显著高于对照组。“自杀式”DNA疫苗是一种基于病毒复制子开发的新型DNA疫苗。Sun等利用PRRSV GP5和CSFV E2基因构建了“自杀式”DNA疫苗pSFV1CS-E2。结果表明,“自杀式”DNA疫苗pSFV1CS-E2共表达GP5和E2,可诱导对PRRSV和CSFV的特异性体液和细胞介导的免疫应答。Jiang等通过构建GP5m和PRRSV M蛋白共表达的“自杀式”DNA疫苗,即PSFV-ORF5m/ORF6,并报道了与PSFV-ORF5m/ORF6相关的典型免疫反应。本实验也为PRRSV DNA疫苗的研制提供了新的思路和途径。与传统疫苗相比,DNA疫苗更安全,制备过程更简单;虽然生产周期短,但基于DNA的疫苗与其他方法具有相同的缺点,如免疫原性低,不能充分解决PRRSV的异质性。“自杀式”DNA疫苗因其安全、高效和易于制备,是DNA疫苗发展的重大突破。蛋白质的低表达可归因于抗原呈递细胞对DNA疫苗质粒的摄取不足。研究人员已经探索了递送DNA疫苗的替代途径和方法,以增强其吸收。皮下注射针已被研究作为通常的皮下和肌肉注射途径的替代。其他方法,如透皮、粘膜和微针应用,以及喷射注射和电穿孔,也已经过测试,以提高DNA疫苗的免疫原性。
7、基因缺失疫苗
基因缺失疫苗是利用基因工程技术切除毒株的毒力相关基因而构建的。Leng等获得了PRRSV基因缺失疫苗株TJM-F92,可在猪的任何生长期接种,保护率为80%。Xu等通过删除HuN4-F112减毒疫苗NSP2区的75个核苷酸,构建了感染性cDNA克隆。将编码新城疫病毒核蛋白(NP)的免疫显性B细胞表位基因片段插入缺失位点。用重组PRRSV免疫仔猪,然后攻毒,接种仔猪产生NP和PRRSV特异性抗体,但没有NSP2抗体。实验组免疫仔猪在攻毒后无临床体征,对照组免疫仔猪在攻毒后10 d全部死亡。结果表明,重组PRRSV rHuN4-F112-∆508-532可作为PRRS的标记疫苗。Hu等构建了带有CMV启动子的日本脑炎病毒(JEV)活疫苗SA14-14-2,并基于JEV DNA将PRRSV GP5/M插入复制子的缺失位点,开发出Balb/c嵌合复制子候选疫苗pJEVREP-G-2A-M-IRES和pJEV-REP-G-2A-M用于动物免疫检测。ELISA数据分析显示,GP5/M复制子诱导较好的免疫应答;因此,基于GP5/M JEV DNA的复制子可以进一步开发成为一种新的、安全的PRRSV候选疫苗。Zhou等对2012- 2016年收集的61个PRRSV NSP2高变区(NSP2HVs)和ORF5进行了测序,除典型的30个氨基酸不连续缺失外,PRRSV中还检测到5个NSP2HVs缺失;因此,本研究提出了一种可用于PRRSV基因缺失疫苗开发的方法。基因删除疫苗通过选择性地删除或突变与毒力相关的基因,将高传染性病毒修饰为弱病毒。这种改变确保疫苗保持其免疫原性,同时防止病毒在动物中轻易恢复其毒力。此外,它允许明确区分野生病毒的自然感染和通过疫苗接种获得的免疫。目前,基因缺失疫苗被广泛用作PRRSV疫苗,相应的开发技术也将有助于新型PRRSV疫苗的研发。
8、合成肽疫苗
合成肽疫苗是以病毒基因组序列衍生的病毒蛋白氨基酸序列为基础,采用化学合成技术人工合成的疫苗。它们通常通过与佐剂偶联来制备。Mokhtar等利用合成全蛋白质组的肽库,对免疫PRRSV-1Olot/91的小群猪进行反复检测和感染,较好地证明了T细胞对PRRSV-1的应答。然而,蛋白质组全结构域肽库的筛选方法和鉴定的抗原需要在下一代疫苗开发的背景下进一步评估。Mokhtar等发现M和NSP5蛋白是多功能CD8和CD4T细胞的保守靶点。然后,通过掺入合成的多聚TLR2/TLR7兴奋剂,将代表M和NSP5的肽包被在疏水修饰的壳聚糖颗粒中,并与含有模型B细胞PRRSV抗原的佐剂一起使用。结果显示CD8 T细胞对M/ NSP5特异性IFN-γ有显著应答;然而,未来的工作应侧重于增强M/NSP5与CD8 T细胞的交叉呈递。在合成肽疫苗的开发中,我们应着重加强M/NSP5与CD8 T细胞的交叉呈递。
合成肽疫苗作为一种新型疫苗,起步较晚,但具有良好的安全性、稳定性和较低的生产成本。这有助于根据疫情快速开发出有效的靶向疫苗,具有良好的发展前景。然而,合成肽疫苗是通过人工合成的方式生产的,单个线性抗原表位在猪体内的保护能力可能不太好,免疫原性较低。因此,串联多个中和性表位并添加合适的佐剂是目前增强合成肽疫苗免疫原性的有效手段,深入研究猪PRRSV与免疫系统的空间构象是进一步开发合成肽疫苗的关键。
9、VLP疫苗
VLP是由一种或几种体外表达的病毒结构蛋白组装而成的中空蛋白颗粒。它们的外观与天然病毒颗粒相似,但它们不含核酸,因此不能繁殖,也没有致病性。它们直径约为20 - 150nm,具有良好的免疫原性,可以在低免疫诱导剂量下使用。免疫后可诱导体液免疫、细胞免疫和粘膜免疫。
VLP疫苗已被商业化开发用于预防和控制其他疾病,临床效果良好。研究人员还探索了PRRSV VLP疫苗的开发并取得了具体进展。先前的研究表明,PRRSV VLPs的形成需要两种以上的蛋白。采用杆状病毒表达系统制备的含有PRRSV GP5和M蛋白的VLPs的免疫活性与天然蛋白没有差异,用不同剂量的VLPs免疫小鼠均表现出特异性免疫活性。有研究表明GP2b在促进VLPs释放中起重要作用。PRRSV的免疫原性基因也可以嵌入到其他病原体的免疫原性基因中合成VLPs,用于同时防治多种病原体。Wang等制备了与主要免疫原性蛋白SIV和PRRSV嵌合的VLPs。动物免疫试验发现,嵌合VLPs可能是一种安全有效的PRRSV和SIV候选疫苗,并能在体内诱导细胞和体液免疫反应。Kang等通过肌注、饮水、滴鼻等方法制备由PCV2、猪细小病毒和PRRSV主要免疫原性蛋白组成的VLPs,并免疫猪,测试其免疫诱导效果。结果表明,VLPs可以同时刺激猪对三种病原体的体液和细胞免疫反应,在猪的所有阶段都具有保护作用,并且可以用作多种疾病的疫苗。这为开发新疫苗提供了基础。利用大肠杆菌表达系统也表达了乙型肝炎病毒核心蛋白与PRRSV GP5的融合蛋白,该融合蛋白可以独立组装形成VLPs。体外实验表明,VLPs能够抑制PRRSV对细胞的感染,且越高浓度的VLPs抑制效果越好。利用新的杆状病毒表达系统进一步制备了含有PRRSV GP5-GP4-GP3-GP2a-M蛋白的VLPs。将PLGA纳米颗粒包埋后,通过鼻内滴注免疫猪,然后激发检测对免疫的影响。结果表明,免疫后猪体内同时诱导体液免疫和细胞免疫,可显著降低猪肺中的病毒载量。然而,关于PRRS VLP疫苗的研究才刚刚开始。PRRSV中能够促进VLPs形成并保持良好免疫原性的蛋白和能够提高免疫诱导效果的佐剂有待进一步探索。
10、潜在的有效疫苗
佐剂对于灭活疫苗的免疫调节作用至关重要。常用的佐剂主要有不溶性铝盐、矿物油佐剂、蜂胶、微生物及其代谢物、免疫刺激复合物、细胞因子等。使用纳米颗粒包封的PRRSV灭活疫苗和聚乳酸-乙醇酸或结核分枝杆菌全细胞裂解液作为佐剂的鼻内注射策略已被广泛用于诱导针对异种PRRSV毒株的交叉保护性抗PRRSV免疫。此外,由大肠杆菌和人参皂苷Rg1的耐热性肠毒素B亚基组成的鼻内佐剂加入灭活的PRRSV疫苗中,可以上调IFN-I信号转导,增强小鼠的耐药反应;然而,还需要进一步的研究来评估这种联合佐剂对猪的免疫保护效果。toll样受体(TLR)激动剂9可减少疫苗接种后的病毒血症并增强非抗原特异性IFN-γ水平。因此,它有望成为与灭活疫苗联合使用的一种有前景的佐剂。Chen等采用新型PRRSV特异性IgM单克隆抗体(Mab)-PR5nf1作为疫苗佐剂,将灭活的PRRSV(KIV)和Mab-PR5nf1与标准佐剂混合制备,增强PRRSV-KIV疫苗介导的保护作用,并将其与普通的KIV和MLV疫苗对免疫系统的作用进行比较。结果表明,在PRRSV-KIV疫苗中添加PRRSV特异性IgM,血清IFN-γ定量和IFN-γ ELISpot检测显示,PRRSV-KIV疫苗可显著提高总生存率和CMI。因此,这种独特的配方与新佐剂结合,可能会增强对PRRSV灭活疫苗的免疫应答。最近的研究也表明,猪肠道菌群可以减少接种灭活疫苗时的应激反应,这可能引导开发新的策略来提高疫苗的效力。在他们的研究中,Maragkakis等人重点研究了接种PRRSV MLV的ID和IM疫苗对Fas相关细胞凋亡的影响。结果显示,PRRSV病毒载量与Fas水平存在显著正相关和中度相关。接种猪血清Fas水平升高提示细胞凋亡受到抑制。这项研究从不同的角度揭示了MLV疫苗的潜在作用。
最优免疫方案不仅与免疫剂量和加强免疫间隔有关,而且疫苗接种途径也是重要因素。常见的疫苗接种方法有肌肉注射、皮内注射和皮下注射。不同接种途径诱导的免疫应答强度也各不相同。肌肉组织只含有少量的常驻免疫细胞。然而,肌肉注射疫苗会导致免疫细胞的大量募集和局部炎症反应。皮肤富含大量具有免疫功能的细胞,参与炎症、修复和免疫反应的初始阶段。皮下组织主要由脂肪细胞组成,只有少量巨噬细胞和淋巴细胞。最近的研究表明,皮内注射可能会降低IL-10和IFN-γ-SC的水平,从而提高疫苗的疗效。肠道菌群对PRRS-MLV疫苗的有效性起着至关重要的作用,为提高MLV疫苗的有效性提供了新的思路。
为了解决现有疫苗免疫原性差的问题,研究人员找到了提高疫苗效力的方法。VCSL1-GP5-N33D是利用反向遗传技术开发的低糖基化嵌合病毒,含有韩国家谱-1野生型株的GP5外区。在发生PRRSV流行的猪场应用vCSL1-GP5-N33D灭活疫苗可获得较高的血清病毒中和抗体滴度,但阴性对照中没有猪出现SVN抗体滴度。动物实验的总体结果表明,vCSL1-GP5-N33D灭活疫苗是一种有希望的候选疫苗。Cui等针对GP5-Mosaic开发了GP5-Mosaic DNA Vaccine Prime/GP5-Mosaic rVaccine,实验结果表明,接种基于GP5-Mosaic的疫苗可导致VR2332和MN184C PRRSV株产生细胞反应性和更高水平的NAs。此外,在MN184C或VR2332攻击后,血清、组织、PAMS和支气管肺泡灌洗液中的病毒载量显著降低,肺部病变较轻。结果表明,采用DNA引物/VACV增强方案的GP5-Mosaic疫苗可以保护猪免受异源病毒的感染。
11、总结与展望
PRRS是全球养猪业最严重的疾病之一,它可以损害猪的免疫器官,引起免疫抑制,并增加对其他疾病的易感性。PRRSV与ADE现象相关,容易发生突变。不同的病毒株需要更好的交叉保护,这使得开发传统疫苗具有挑战性。目前的灭活疫苗具有免疫诱导作用持续时间短、接种剂量大、成本高的特点。此外,MLVs的毒力也有增加的风险。此外,很难确定抗体是在应对野生病毒感染或疫苗免疫时产生的。DNA、亚单位或病毒载体疫苗也已进行了测试,但它们作为PRRSV-MLV替代品的潜在价值仍不确定(表3)。商业化的VLP疫苗产品很少,还有许多问题需要解决,如基因序列的选择和优化、表达系统的选择、表达量的提高、VLP的大规模纯化、成本的降低、佐剂的选择和添加比例等。以及疫苗功效的评价。因此,迫切需要开发一种新的疫苗来预防和控制这种疾病。此外,在系统分析的基础上,可以利用DNA重组或合成PRRSV结构蛋白基因片段来开发新的PRRSV杂交毒株。此外,如果NAs的保守表位能够被识别和表征,则可以在设计中考虑这种新的杂交基因诱导针对异种PRRSV的交叉保护的能力。可被期待的是,结合先进技术设计的PRRSV疫苗将比现有疫苗表现出更好的有效性和安全性。总之,本文建议在PRRSV疫苗开发领域需要继续进行研究和开发,以解决现有疫苗的局限性,并提供更好的PRRSV保护。
表3.几种PRRSV疫苗的比较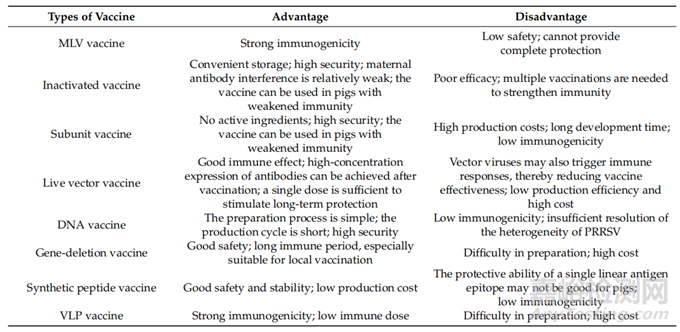
参考资料:Zhang H, Luo Q, He Y, Zheng Y, Sha H, Li G, Kong W, Liao J, Zhao M. Research Progress on the Development of Porcine Reproductive and Respiratory Syndrome Vaccines. Vet Sci. 2023 Jul 28;10(8):491. doi: 10.3390/vetsci10080491. PMID: 37624278; PMCID: PMC10459618.



